
The following points highlight the five main fundamental concepts related to energy. The concepts are: 1. Solar Radiation 2. Concept of Productivity 3. Concept of Energy Subsidy 4. Concept of Source-Sink Energetics 5. Energy beyond the Producers.
1. Solar Radiation:
The sun has a temperature and composition such that hydrogen is transmuted to helium with a concomitant release of considerable energy in the form of electromagnetic wave. About one fifty-millionth of the sun’s tremendous energy output reaches the earth’s outer atmosphere and it does so at a constant rate.
This constant is referred to as solar flux (solar flux is the amount of radiant energy of all wavelengths that cross a unit area or surface per unit of time). The flux at any given point varies seasonally with latitude, and also diurnally.
This flux gets depleted as it passes through the trophosphere and only about 48 per cent reaches the earth surface (Fig. 4.1). The extraterrestrial sunlight reaches the ionosphere at a constant rate of 2 g cal/cm2/min and is referred to as solar constant. Due to dilution and the dispersed nature of light, only a small fraction (at most 5 percent) of visible light can be converted into organic matter by the producers of the ecosystem.

2. Concept of Productivity:
Primary productivity of an ecological system is the rate at which radiant energy of the sun is converted into organic substances by the photosynthetic and chemosynthetic activities of producers.
The following are some of the important terms used in the production process:
1. Gross primary productivity (GPP):
It is the total rate of photosynthesis which includes the energy used-up in respiration (R) during the measurement period. It is also referred to as total photosynthesis.
ADVERTISEMENTS:
2. Net primary productivity (NPP):
It is the rate of storage of organic matter in the tissues of the producers that exceeds the respiratory (R) use. It is also termed as net assimilation.
To estimate the GPP the amount of plant respiration is added to the NPP:
GPP = NPP + R
ADVERTISEMENTS:
3. Net community productivity:
It is the rate of storage of organic matter not used by heterotrophs, or, in other words, it is NPP minus heterotrophic grazing (consumption).
4. Secondary productivity:
It is the rate of energy storage at consumer levels. Secondary productivity cannot be divided into gross and net amounts as the consumers use only food materials already produced. The total energy flow at heterotrophic levels should be designated as assimilation and not production.
5. Net ecosystem production:
As there is intimate relationship between terrestrial plants and the soil, ecologists, when measuring NPP, often consider the two as a unit. The net ecosystem production (NEP) is the difference between GPP and the energetic costs of both the plants and the soil organisms.
NEP = Gross primary production – respiration of plants – respiration of soil organisms.
3. Concept of Energy Subsidy:
In natural and cultivated ecosystems, high rates of primary production occur when physical factors (water, nutrients etc.) are favourable and when auxiliary energy from outside the system enhances growth or rates of reproduction within the ecosystem. Such secondary energy supplements are termed auxiliary energy flow or energy subsidy.
Some examples are wind and rain in a rainforest, tidal energy in an estuary, fossil fuel or fertilizers and animal or human work energy used in cultivating a crop. While evaluating the productivity of an ecosystem, one must consider not only the energy subsidies, but also the energy drains resulting from harvest, pollution, climatic and other stresses that drain away energy from the ecosystem.
4. Concept of Source-Sink Energetics:
ADVERTISEMENTS:
When organic production by one ecosystem (source) is exported to another less productive ecosystem (sink), the concept is termed as source-sink energetics. A productive estuary may export organic matter or organisms to coastal waters that are less productive. Therefore, the productivity of an ecosystem is determined not only by the rate of production within it but also the imports or that exported from a source system.
Energy fixation and estimates of Primary Production:
The autotrophs convert radiant energy into chemical energy. The measurement of the amount and rate of energy fixation is based on the photosynthetic equation:
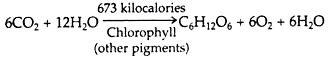
Edgar Transeau (1926) estimated the accumulation of energy by a mid-western cornfield in a single growing season. He calculated that there were 2,675 kg of Carbon in the 10,000 corn plants harvested. Since this carbon entered the plants only through photosynthesis, it would be equivalent to 6,687 kg of glucose:
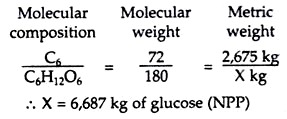
Transeau then obtained the total or gross production (GPP) of 8,732 kg by adding to the above net production (6,687 kg) an equivalent amount of glucose (2,045 kg) that he estimated had been used in cellular respiration during the growing season.
Since 3,760 kilocalories are required to produce one kilogram of glucose; 33 million kcal would have been incorporated in gross production of which 7.7 million kcal was used in metabolic activities. As the total solar energy available to the corn field was known, it was possible to calculate the efficiency of energy utilised, which was as low as 1.6 per cent.
![]()
![]()
Further calculations revealed that in the case of the cornfield, the energy loss by respiratory activity is 23.4 percent:
![]()
= ![]()
Thus it is interesting to note that although relatively little of the total available energy was utilised in photosynthesis, the plants were quite efficient in converting 76.6 percent of the captured energy to biomass.
In another study of a two-year period, conducted by Jana and Kundu (1991) in two stocking ponds at Kalyani, W. Bengal, it was found that the photosynthetic efficiency of phytoplankton ranged from 1.18 to 1.28 percent. Kormondy (1986) stated that the efficiency of energy capture in gross production under natural conditions was seldom more than 3 percent.
The annual mean of primary production conversion efficiencies and ratio, in the two stocking ponds is in Tables 4.2 and 4.3. In these two ponds, due to the presence of grass carp, macrophytes in the form of Hydrilla sp. was added to the system resulting in Allochthonous source of energy.


Generally, the photosynthetic efficiency in aquatic ecosystems is a bit lower due to reduced light penetration in aquatic medium. Open oceans and deserts also are much less productive due to nutrient limitation.
Energy expenditure of producers in self-maintenance and growth, and loss via respiration also varies considerably from 15 to 24 percent. However, in temperate forests, this expenditure reaches 50 to 60 percent and 70 and 75 percent in tropical forests.
H. T. Odum (1956) has proposed a classification of ecosystem based on the ratio of photosynthesis (P) to respiration (R). According to this scheme, a stabilized system is one in which the ratio of P/R = 1; a system in which P exceeds R (P/R > 1) is considered autotrophic (Example: fertile agriculture etc.) and one in which R exceeds P (P/R < 1) is said to be heterotropic (Example: polluted streams, swamp waters etc.). Fig. 4.2 shows the position of various community types in the above classification.

5. Energy beyond the Producers:
In the food-chain, the herbivores feed upon the autotrophs or producers. The entire gross energy produced by the autotrophs are not transferred to the herbivores, rather, only about 15-20 percent or even less is transferred, as some is lost due to respiration and decay of autotrophs and a major amount remains unutilized. Similar is the case in the carnivores which feed upon the herbivores.
The fate of energy flow in the ecosystem can be best described by taking the experiment conducted by Raymond Lindeman (1942) in the Cedar Bog Lake, Minnesota. He estimated the gross energy production by the autotrophs to be 111 gcal cm−2yr−1 with an efficiency of solar energy capture of 0.1 percent. 21 percent of the gross energy production or 23 gcal cm−2yr1 was used up by the autotrophs in the biological reactions occurring at the cellular level.
Of the rest, only 15 gcal cm−2yr−1 was consumed by the herbivores. Decomposition accounted for about 3.4 percent (3 gcal cm−2yr−1) of net production and the remainder, 70 gcal cm−2y−1, was not utilised at all. It is obvious that much more energy was available for the herbivores than was consumed and that the said ecosystem was especially prodigal in energy utilisation beyond the autotrophic level.
Of the total energy incorporated at the level of the herbivore, namely 15 gcal cm−2yr−1, 30 percent or 4.5 gcal cm−2yr−1 is used in metabolic activity. There is then considerable more energy lost via respiration by herbivores (30 percent) than by autotrophs (21 percent).
Again, there is considerable energy available for the carnivores, namely 10.5 gcal cm−2yr−1 or 70 percent, which is not entirely utilised, in fact, only 3.0 gcal cm−2yr−1 or 28.6 percent of net production passes to the carnivores. This is more efficient utilisation of resources than what occurs at the autotroph → herbivore transfer level, but, nonetheless, is still quite profligate.
At the level of the carnivore, about 60 percent of the carnivores intake is consumed in metabolic activity and the remainder becomes part of the non-utilised sediments. It is interesting to note that there is high respiratory loss in the carnivores (60 percent) in comparison with herbivores (30 percent) and autotrophs (21 percent) in this ecosystem. This is mainly due to the greater locomotory activity of carnivores for which they have to spend considerable amount of energy than others.