
During the growth of the oocyte not only does the amount of cytoplasm increase in quantity, but it changes in quality by the elaboration and regular distribution of various cell inclusions and specially modified parts of the cytoplasm which, are essential for the development of the embryo.
In most animals which have been investigated with modern methods, young oocytes show a very similar and fairly simple organization. The cytoplasm in young oocytes is rather poor in complex inclusions and possesses practically none of the specialized structures found in the adult oocyte and mature egg.
Large amounts of ribonucleic acid are present in the cytoplasm of young oocytes. As seen with the electron microscope the cytoplasm is finely granular. The granules are almost certainly ribonucleoprotein in nature, as the cytoplasm is strongly and uniformly stained by pyronin or azure B, stains used to reveal ribonucleic acid.
Mitochondria are fairly scarce in young oocytes but may increase in numbers considerably during the growth of oocytes and may in some animals (amphibia, birds), be aggregated in the form of large “mitochrondrial clouds”.
ADVERTISEMENTS:
Because mitochondria are carriers of oxidative enzymes, the overall oxygen consumption increases during the growth of the oocyte. Young frog oocytes, before the beginning of yolk formation, have been found to absorb 0.69 cu. mm. of oxygen per cubic millimeter of oocyte. At the stage when the yolk platelets begin to form, the oxygen consumption rises to 1.5 cu. mm. per cubic millimeter.
In older oocytes nearing maturity, the oxygen consumption per cubic millimeter falls to 1.2 cu. mm. This is still a high value, considering that a large part of the volume of the oocyte is filled with inert yolk; thus, the remaining active cytoplasm continues to respire at a high rate. Golgi bodies are found in younger oocytes around the centrosome.
In oocytes of some mammals, Golgi-type membranes form a large spherical mass which later disperses. In oocytes which have advanced in growth, small Golgi bodies become scattered throughout the whole cytoplasm, or they may be located in the subcortical cytoplasm, sometimes distributed there at fairly regular intervals. In mature oocytes Golgi bodies sometimes disappear completely, possibly indicating that these organelles have been used up or transformed into some other structures.
Typical endoplasmic reticulum, in the form of double lamellae with ribosomes attached to their outer surfaces, is very rarely found in young oocytes. The cytoplasm instead contains fairly numerous vesicles surrounded by a simple membrane. These vesicles are generally believed to be equivalent to or a modification of the endoplasmic reticulum.
ADVERTISEMENTS:
Ribosomes are often attached to the surface of the vesicles. A different kind of membranous structure is, however, often found in oocytes; it takes the form of stacks of double membranes, sometimes in parallel, and sometimes in spiral arrangement.
The membranes usually do not have ribosomes attached to them but instead are perforated by pores, which closely resemble the pores of the nuclear membrane. For this reason the membranes are regarded by some authors as being derived from the nuclear membrane.
Yolk appears in the oocytes in the second period of their growth. Accordingly, many embryologists distinguish two periods of oocyte development – the previtello-genesis period and the vitellogenesis period. Growth is very much accelerated during the vitellogenesis period; this period is thus also a period of rapid growth.
The most usual form of food storage in the egg consists of granules of yolk. Yolk is not a definite chemical substance, but rather a morphological term; the chemical substance may not be the same in all cases. The main chemical components of yolk are proteins, phospholipids, and to a lesser extent, neutral fats.
ADVERTISEMENTS:
Depending on which components predominate, we can distinguish “protein yolk,” which may contain variable amounts of lipids as well as protein, and “fatty yolk,” which in addition to phospholipid and fat may contain some admixture of protein. Protein yolk and fatty yolk are present side by side in the eggs of many animals.
Protein yolk is the main form of food reserve in many invertebrates (for instance, in echinoderms) and in lower chordates (Amphioxus, tunicates). The amount of yolk in these animals is relatively small; in the sea urchin Arbacia the yolk granules take up about 27 per cent of the total volume of the egg. The yolk granules are fine and fairly evenly distributed in the cytoplasm of the egg. Eggs with a small amount of yolk are called oligolecithal.
In amphibian eggs the protein yolk is found in the form of rather large granules, usually described as the yolk platelets. The yolk platelets have an oval shape and are flattened in one plane. The cytoplasm is densely packed with them.
The amphibian egg is much larger, so that not only the relative but also the absolute amount of yolk is far in excess of that found in the eggs of Amphioxus or the echinoderms. The distribution of yolk in the amphibian egg is distinctly unequal – the yolk platelets are densest in the lower part of the egg, and there is relatively more cytoplasm in the upper part of the egg. Eggs of this type are known as telolecithal.
The yolk platelets of amphibians contain two main proteinaceous substances – phosvitin and lipovitellin. Phosvitin is a highly phosphorylated protein (phosphorus content – 8.4 per cent), with a molecular weight of 35,000. Lipovitellin is a protein with a very much larger molecule (molecular weight 400,000) and containing a very considerable amount of bound lipid (17.5 per cent).
In the yolk platelets two molecules of phosvitin are associated with each molecule of lipovitellin in a structural unit, the units being arranged in the platelet in a crystalline lattice with regular hexagonal packing. This can be clearly seen in electron micrographs of amphibian yolk.
In addition to the yolk platelets the amphibian egg contains stored supplies in the form of lipid and glycogen. Lipid is distributed throughout the cytoplasm of the egg in the form of organized inclusions, the lipochondria, which consist of an internal core of lipid surrounded by a thin protein coat.
The lipochondria are much smaller than the yolk platelets and are roughly spherical in shape. Glycogen is present in the egg cytoplasm in the form of small granules. Fat may also be stored in the egg for the nourishment of the developing embryo, but its quantity is relatively small in the lower vertebrates.
In a mature amphibian egg, protein yolk constitutes roughly 45 per cent of the dry weight, lipids 25 per cent, and glycogen 8.1 per cent. Only about 20 per cent of the dry weight of the mature egg is active cytoplasm.
ADVERTISEMENTS:
Cyclostomes, elasmobranchs, ganoids, and the lungfishes have eggs with a distribution of food reserves much the same as in amphibians, though the amount of yolk may be greater, especially in some selachians.
In the higher teleosts, on the other hand, the yolk becomes segregated from the cytoplasm and concentrated in the interior of the egg, while most of the cytoplasm is found in a thin surface layer covering the yolk and thickened on the uppermost side of the egg in the shape of a cytoplasmic cap. The nucleus of the egg in this case lies inside this cytoplasmic cap. The core of the yolk platelets of cyclostomes and fishes, as in amphibians, has a crystalline structure.
In the bony fishes, fat may be present in the form of large fat droplets inside the mass of yolk. The number and size of the fat droplets is typical for different families of fishes. In some species only one large droplet is present, while in others there are a large number of smaller droplets.
The yolk of a bird’s or a reptile’s egg, as in the case of the bony fishes, lies in a compact mass in the interior of the egg, and the cytoplasm is restricted to a thin layer on the surface, with a thickened cap of cytoplasm on the upper side. As in bony fishes, this cytoplasmic cap also contains the nucleus of the egg. Most of the yolk is liquid, but about 23 per cent is in the form of solid “yolk spheres.”
The yolk as a whole contains 48.7 per cent water, 16.6 per cent proteins, 32.6 per cent phospholipids and fats, and 1 per cent carbohydrates. The proteins of the avian yolk are chemically closely related to those of the amphibian yolk – phosvitin and lipovitellin are the main components.
Part of the lipovitellin is bound to phosvitin; this part of the yolk protein is insoluble in water. The remainder of the lipovitellin is water soluble. The fatty portion of avian yolk is predominantly neutral fat (50 per cent of the yolk dry weight), the rest being phosphatides and cholesterol.
Some of the invertebrates also have developed eggs in which the relative amount of yolk is high and more or less segregated from the cytoplasm. In cephalopods and some gastropods among the molluscs the eggs are telolecithal, much as in the lower vertebrates.
Arthropods, especially insects, have developed a different type of egg – the yolk is concentrated in the interior of the egg, and the cytoplasm is distributed as a thin coat on the external surface; however, there is also an island of cytoplasm in the center of the egg. This island, surrounded on all sides by yolk, contains the nucleus of the egg cell. Eggs of this type are called centrolecithal.
For a long time it has been believed that the production of yolk in the oocyte is the result of the activity of a special body, found in the oocytes of many animals, which accordingly was called the “yolk nucleus” or “yolk nucleus of Balbiani.”
This body was first discovered in the oocytes of spiders, but later it was shown to be present in avian oocytes and in the oocytes of some mammals and amphibians. In the early stages this body lies next to the nucleus, but in later stages it breaks up and its fragments are distributed to the periphery of the oocyte.
As it is at the periphery of the oocyte that the yolk platelets appear first, the conclusion has been drawn that vitellogenesis (production of yolk) is initiated by the components of the “yolk nucleus.” At present, it is not possible to support this conclusion, at least not in a general form. Electron microscopic investigations have shown that “yolk nuclei” in different animals are not always of a similar nature.
In spiders the yolk nuclei consist of numerous stacked membranes with mitochondria and vesicular structures surrounded by these membranes or interspersed between them. There is no indication that these bodies have anything to do with the formation of yolk.
In amphibians the name “yolk nucleus” has been applied to the mitochondrial cloud, and the same appears to be true of the avian oocyte, although an electron microscopic verification of the nature of the body in the bird is still lacking. In amphibians (Xenopus), it was found that although parts of the mitochondrial cloud reach the periphery of the oocyte, they do so too late to participate in yolk platelet formation which starts before the breakup of the mitochondrial cloud.
In mammals a spherical body lying in the cytoplasm next to the nucleus and having, in the living state, the same appearance as the mitochondrial cloud of the amphibian oocyte, consists of a mass of Golgi-type membranes and is now referred to as the Golgi material. The Golgi material in mammals cannot be involved in yolk formation for the simple reason that, in the oocytes of higher mammals, yolk, at least in the form of yolk platelets, is completely lacking.
Apart from the “yolk nucleus,” several cellular organoids have been considered responsible for yolk formation in different animals by different authors. The organoids most often mentioned in this connection are the endoplasmic reticulum, the Golgi bodies, and the mitochondria.
The evidence is rather contradictory. The discrepancies may perhaps be resolved if one considers the actual site of synthesis of the yolk proteins. It has been indicated previously that the yolk is not necessarily synthesized in the oocytes, but may be produced elsewhere in the body (in the liver in the case of vertebrates) and is then transported in a soluble form via the blood stream and the follicle cells and is re-deposited in the oocyte.
In other cases no such transport of ready-made yolk has been recorded, and it seems plausible that the yolk is actually synthesized from simple components in the oocyte by the usual process of protein synthesis, that is, on the ribosomes of the oocyte itself.
This last process very likely occurs in animals such as coelenterates, which do not have a circulatory system. In a medusa the first visible yolk particles have been found in cisternae of the Golgi bodies, but it is plausible that the actual synthesis occurs on the ribosomes of rough endoplasmic reticulum associated with the Golgi bodies, which serve as the “packaging” organelle, as they do in some secretory cells of adult animals.
Some carbohydrate may be added in the process, the latter synthesized by the Golgi bodies themselves. In crustaceans yolk granules actually appear in the interior of laminar or vesicular endoplasmic reticulum. It has been found also that radioactively labeled leucine is taken up into the granules—a very strong indication that yolk is synthesized locally, directly from amino acids.
While yolk formation in connection with the endoplasmic reticulum and the Golgi bodies conforms to what is generally known about the process of protein synthesis in the cell, this cannot be claimed to be the case where yolk appears in or close to mitochondria of oocytes.
There is conclusive proof that in amphibians and fishes the yolk platelets are formed inside modified mitochondria. In this process the inner mitochondrial membranes become dislodged and eventually become arranged in concentric layers, while the interior of the mitochondrion is taken up by the main body of the yolk platelet. Occasionally a yolk crystal may be seen lying inside a swelling of a mitochondrion, which in other parts still shows a typical structure with systems of transverse cristae.
A somewhat similar origin of protein yolk has been described in gastropod molluscs. The protein yolk platelets, which in gastropods also show crystalline structure, are formed either actually inside mitochondria or in between clumps of mitochondria.
It has been indicated previously that in some animals the yolk is not synthesized in the oocytes at all but is produced elsewhere in the body (in the liver, in the case of vertebrates), and that it is then transported in a soluble form via the bloodstream and the follicle cells to the oocyte, where it is finally deposited in the form of yolk platelets or yolk granules.
In this connection, some properties of the yolk proteins are of particular interest. The yolk protein phosvitin is insoluble in water when it is fully phosphorylated. If, however, part of the phosphate is removed by the action of a phosphatase, the remaining phosvitin becomes water soluble.
The reverse would then also hold true; namely, if soluble partially phosphorylated phosvitin were more fully phosphorylated in the oocyte, it would become insoluble. An enzyme, protein kinase, has actually been found in the frog ovary which is capable of promoting the incorporation of phosphate into partially phosphorylated phosvitin. The phosphate is taken from ATP.
The reaction is fully reversible and takes place according to the following equation:
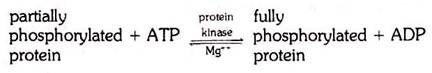
The action of protein kinase is restricted to only one component of the amphibian yolk (phosvitin). In the yolk crystal, however, the lipovitellin is bound to phosvitin, and therefore the rendering of phosvitin insoluble fixes the lipovitellin in position as well.
Now it is very important that protein kinase is a mitochondrial enzyme, as this explains why the yolk platelets may be formed inside or in connection with mitochondria – it is there that the yolk proteins are converted from the soluble into the insoluble form. Obviously, much remains to be done to explain the deposition of yolk in such animals as birds, and also to clarify the origin of fatty yolk.