
The below mentioned article provides a study note on the Genetic Linkage:- 1. Discovery of Genetic Linkage 2. Types of Genetic Linkage 3. Linkage Group.
Discovery of Genetic Linkage:
In 1905, William Bateson, E. R. Saunders and R. C. Punnett discovered the exception to the law of independent assortment of gene pairs while investigating the inheritance patterns for various traits in the sweet pea.
In one study, they examined the simultaneous inheritance of flower colour, dominant purple (P) versus recessive red (p), and pollen shape, dominant long (L) versus recessive round (l). They carried out a di-hybrid cross as Mendel had done for traits in the garden pea. They crossed PP LL with pp 11 and self-fertilized the F1 hybrids (Pp LI). They counted the numbers of different types of F2 plants.
But instead of the expected 9:3:3:1 ratio under independent assortment, they obtained the results as given below:

They observed that the two parental phenotypes were in excess in the F2 progeny. They predicted that there might be a physical connection between the parental alleles — the dominant alleles from one parent and the recessive alleles from the other parent. This phenomenon they termed as coupling — a situation where the dominant genes seemed to ‘stick together’.
On the other hand, they attributed the low numbers of the F2 types, those having a dominant phenotype at one gene and a recessive at the other, to some innate negative affinity of dominant and recessive alleles, which they termed repulsion — a situation where non-allelic dominant genes were ‘repelled’ from each other.
However, Bateson, Saunders and Punnett could not explain the discrepancy that occurred in their observed results from that expected by Mendel’s second law. They had to await till the development of Drosophila as a genetic tool.
The proper explanation of Bateson’s observation came later from the research of T. H. Morgan who used two genes in Drosophila for his study. One gene was for eye colour (recessive purple, pr, and dominant wild type red, pr+, alleles) and the other gene affected wing development (recessive vestigial, vg, and dominant normal, vg+ alleles).
ADVERTISEMENTS:
Instead of examining the F2 progeny of a di-hybrid cross, Morgan used a test cross for both genes simultaneously. First, he made cross between flies with purple eye colour (pr) and vestigial wing (vg), and flies with red eye (pr+) and normal wing (vg+). The resulting heterozygous F, females (pr pr+ vg vg+) were then test crossed:

These results are a drastic deviation from Mendel’s prediction of 1 : 1 : 1 : 1 ratio, and these indicate a coupling of genes. Although the four gametic numbers were not those expected from independent assortment.
The two parental gametes pr+ vg+ and pr vg, were nearly equal in number, and the two non- parental gametes, pr+ vg and pr vg+, were also nearly equal in number, but much less ‘frequent than the parental gametes.
ADVERTISEMENTS:
The gametes either with two wild-type or two mutant alleles are called coupling gametes. The gametes with one wild-type and one mutant allele are repulsion gametes. In other words, the two parental gametes were in coupling and the two non-parental gametes were in repulsion.
In a second experiment, Morgan crossed two different genotypes, each of which was homozygous for a wild-type allele at one gene locus and homozygous for a recessive allele at another gene locus (pr+ pr+ vg vg x pr pr vg+ vg+). The F1 (female) flies, double heterozygotes, were then crossed to the tester line pr pr vg vg (male).
The results of this test cross were as follows:

The results were quite different from the first cross. In this case, the progeny numbers of the single dominants were much larger than expected, while those of the double dominants and double recessives were much lower than expected. In other words, there were more repulsion types than coupling types.
To explain his observations, Morgan suggested that the genes having the alleles for these traits are on the same chromosome. As a result, alleles at these genes tend to remain associated between generations because they are physically linked to each other.
However, instead of these chromosomes always staying intact during the production of gametes (meiosis), in some chromosomes a physical exchange of non-sister chromatids occurs (crossing over or recombination) between the homologous chromosomes. As a consequence, gametes with new gene combinations result.
The intact chromosomes with original gene arrangements are called parental chromosomal type or non-recombinant, while the new combinations are called cross-over type or non-parental chromosomal type or simply the recombinant.
The general conclusions derived from the above mentioned experiment were:
ADVERTISEMENTS:
Genes that are on the same chromosome are said to be linked, and the general phenomenon that the genes lying on the same chromosome try to be inherited together from generation to generation as an inseparable unit, is called linkage.
Thus, the genes that try to show linkage or in other words, the genes that lie on the same chromosome, tend to be inherited together. It should be remembered that such genes are referred to as ‘linked” rather than “coupled”.
Actually coupling and repulsion have come to indicate two different types of linkage conformation in a double heterozygote, as follows:

In other words, coupling refers to the linkage of two dominant or two recessive genes, whereas repulsion indicates linkage of one dominant and one recessive gene in a double heterozygote.
Types of Genetic Linkage:
According to the degree, linkage may be classified into complete and incomplete types. Genes lying on the same chromosome might always be transmitted together and these genes might not undergo independent assortment. Linkage between these genes is considered as complete. Complete linkage between genes on the same chromosome is rare in most sexually reproducing species.
However, in Drosophila, there is complete absence of crossing-over in males; so in males there is 100% linkage in different genes of the same chromosome. Gene pairs in most linkage groups assort at least partially, independent of each other and these linked genes are not always transmitted together. Linkage between these genes is considered as incomplete.
In Drosophila, gray body colour and long wing are two dominant linked characters; the genes for these characters may be indicated as b+ and v+ and their location on the chromosome may be indicated as b+ v+. Again, black body and vestigial wing are two linked recessive characters.
These genes may be indicated as b and v and their location on chromosome as b v. Pure gray long fly is crossed with pure black vestigial fly. The F1 hybrids are phenotypically gray long and their genotype will be ![]() . Now a male heterozygote fly is test crossed with a double recessive female. The test-cross offspring were gray long and black vestigial in the ratio of 1 : 1. In this case it is seen that two linked dominant characters gray and long remain together in the P1 flies, F1 hybrid flies and 50% of the test cross offspring’s.
. Now a male heterozygote fly is test crossed with a double recessive female. The test-cross offspring were gray long and black vestigial in the ratio of 1 : 1. In this case it is seen that two linked dominant characters gray and long remain together in the P1 flies, F1 hybrid flies and 50% of the test cross offspring’s.
Again the two recessive linked characters black and vestigial remain together in the P1 black vestigial flies and 50% of the test cross offspring’s. Thus, we get either grey long combination or black vestigial combination for more than one generation and do not get a mixture like grey vestigial or black long in any generation. Thus, this example shows complete linkage between two dominant traits grey and long and in between two recessive traits black and vestigial.
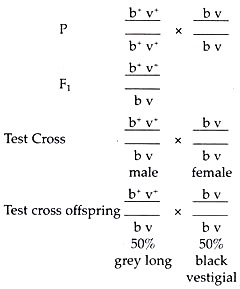
Complete Linkage in Drosophila:
It is important to note that for the test cross the male hybrid Drosophila is test crossed with a female, fly because only in males there is no cross-over and complete linkage can be shown. If the female hybrid is taken, complete linkage cannot be shown as crossing-over occurs in female.
A case of incomplete linkage can be illustrated as follows: pure grey bodied long winged and black bodied vestigial winged were crossed. A female heterozygote grey long fly is crossed with double recessive black vestigial male.
In females, cross-over occurs as a result of which it produces four kind of gametes, two non-cross-over types of gametes are b+ v+ and b v, and two crossover type of gamete are b+ v and b v+. In this case, 82% of the test cross offspring’s are non-cross-over type, of which 41% are grey long and 41% are black vestigial; 18% of the total offspring’s are cross-over type of which 9% grey vestigial and rest 9% black long.
The cross is shown below:

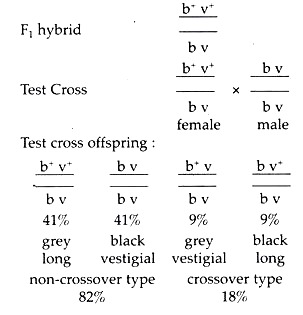
Incomplete Linkage in Drosophila:
In this case, the two dominant characters grey long remain together in P and F1; but after test cross, in addition to the original combination of characters, new combinations like, gray vestigial and black long are appeared i.e., dominant and recessive traits may appear together due to occurrence of crossing-over in the F1 hybrid females.
Thus, it is a case of incomplete linkage. It is important to note that in the test cross, the hybrid fly is female. A male hybrid cannot be selected for the test cross in this experiment, as in Drosophila, there occurs no crossing-over in males.
Genetic Linkage Group:
Genes located on the same chromosome are said to belong in the same linkage group. So each chromosome represents one linkage group. The total number of linkage groups in an organism is equal to the number of chromosome in one haploid set.
In human, the number of linkage groups in female will be 23, 22 for the autosomes and 1 for the X chromosome. Whereas in male there are 24 linkage groups, 22 for autosomes, 1 for X chromosome and 1 for Y chromosome. In Drosophila female, the number of linkage group is 4, 3 for autosomes and 1 for X chromosome and in males there are 5 linkage groups, 3 for autosomes, 1 for X chromosome and 1 for Y chromosome.