
Study Notes on Scypha:- 1. Meaning of Scypha 2. Structure of Scypha 3. Canal System 4. Circulation of water 5. Microscopic Organisation 6. Skeletal System 7. Nutrition 8. Respiration 9. Excretion 10. Nervous System and Response Mechanism 11. Reproduction.
Contents:
- Meaning of Scypha
- Structure of Scypha
- Canal System in Scypha
- Circulation of Water in Scypha
- Microscopic Organisation of Body Wall in Scypha
- Skeletal System in Scypha
- Nutrition in Scypha
- Respiration in Scypha
- Excretion in Scypha
- Nervous System and Response Mechanism in Scypha
- Reproduction in Scypha
1. Meaning of Scypha:
Scypha, formerly called Sycon, exhibits the first stage of body wall folding and is called syconoid sponge type. Grantia is another well-known example of syconoid type.
Here the body wall has become “folded”, forming external pockets extending outwards from the central cavity, the spongocoel. The various species of Scypha are S. ciliatum, S. coronata, S. elegans, S. lingua, S. raphanus, S. gelatinosum etc.
ADVERTISEMENTS:
Etymology:
Greek: Skyphos, cup
Scypha is commonly known as urn sponge due to its urn shape. It is also called crown sponge as the fringe of long and straight spicules looks like a little crown.
Habit and Habitat:
ADVERTISEMENTS:
Scypha is a marine sessile sponge and remains permanently attached to submerged rocks and other solid substrata. It is widely distributed and is especially abundant in places where wave action provides with an adequate food supply and with well oxygenated water. It is a branched, colonial sponge, but solitary individuals are also seen.
2. Structure of Scypha:
Scypha has the structure of solitary or branched cylinders each of about 2-5 to 7-6cm in length and 0-5 to 0-6 cm in diameter. Each resembles a slender vase-like cylinder, slightly bulging in the middle. All the branches are connected together at the base, which remains attached to the substratum.
One or more buds may be found on the side of the body. The cylinders are flexible, though of tolerably firm consistency. They range from white through grey to light brown in colour, depending upon the species and its surroundings.
ADVERTISEMENTS:
Close inspection of the surface reveals the presence of innumerable minute inhalant pores or ostia. Each cylindrical branch has an opening at its free end called osculum. The region below the osculum is somewhat narrow forming a collar region (Fig. 1.21). The osculum is protected by a fringe of mono-axon spicules.

3. Canal System in Scypha:
Scypha, like other sponges, possesses the phylum’s characteristic canal system, which plays a very important role in its life. Its body wall is folded to form regularly arranged alternate invaginations and evaginations, thereby establishing the sycon type of canal system.

The various components of the Sycon type of canal system (Fig. 1.22) in the cylindrical body of Scypha, can be best seen if it is bisected longitudinally. It is observed that the osculum leads into a narrow tubular cavity called the para-gastric cavity or gastral cavity or spongocoel.
Osculum is provided with sphincters to regulate the rate of water flow in the body. Sphincters are lined by special contractile pinacocytes, called myocytes (Greek: myos, muscle; kytos, cell).
The body wall lining the spongocoel is out pushed at regular intervals as finger-like projections called radial canals. The wall of the radial canal is lined with flagellated choanocytes. The radial canal is connected to the spongocoel through small openings called internal ostia or apopyle (Greek: apo, away from; pyle, gate). Apopyles are surrounded by contractile myocytes serving as a sphincter (Fig. 1.23).

In between two successive radial canals, a tubular space called incurrent canal is present, which is lined by pinacocytes. The radial canals and incurrent canals are thus arranged alternately. The wall between the incurrent and radial canals are pierced by numerous minute pores called prosopyles (Greek: pros, near).
ADVERTISEMENTS:
The incurrent canal opens to the exterior through dermal pores or ostia (Latin: ostium, door), but ends blindly at their inner end. The presence of contractile cells or myocytes around the ostia (Fig. 1.24) help in regulating its diameter and thus regulates the amount of water entering into the body of Scypha.

4. Circulation of Water in Scypha:
Circulation of water in Scypha takes place as given below
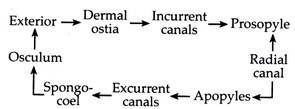
The course of water current has been studied by the application of fine carmine particles to the surface of the body. Water rushes into the body through numerous ostia present along the external surface. Each ostium or dermal pore leads into an incurrent canal.
From here water flows into the radial canal through the prosopyle. Water, from the radial canal passes out through apopyles into the spongocoel. The spongocoel is a common chamber within which all the radial canals of the body opens.
The spongocoel ultimately pushes out the water to the exterior, through the osculum. Therefore, water enters the body through numerous openings (ostia) but passes out through a single opening (osculum). The synchronous beating of the flagella of choanocytes in the radial canal produces a current, which in one hand draws the water inside and on the other hand forces it to go outside.
5. Microscopic Organisation of Body Wall in Scypha:
The structural plan of the body wall of Scypha is focused at a cellular level. The cells do not differentiate into many distinctive types found in higher metazoa. Each cell has quite broad functional capacities. Sponges as a whole are considered to lack well organised tissues.
The microscopic organisation of body wall shows the presence of a single layer of cells covering the external surface of the body, designated as the dermal layer and composed of large pinacocyte cells (Fig. 1.25).
The spongocoel is lined by a layer of flattened endodermal choanocyte cells that are flagellated. The intermediate layer, in between the above two layer of cells, consists of a gelatinous matrix with spicules and numerous amoeboid cells.
i. Pinacocytes:
(Greek: pinako, plank) Pinacocytes (Fig. 1.26A) cover the outer surface and line all internal canals or spaces that are not covered with choanocytes. Pinacocytes sometime form a syncytial epitheloid membrane. Externally lying pinacocytes on the outer body surface are called exo-pinacocytes, while the internal pinacocytes are called endopinacocytes.
The pinacocytes are large, flattened, scale-like cells, closely cemented together by their thin margins. It contains a somewhat centrally placed nucleus. Pinacocytes are highly contractile and can either reduce or increase the surface area of the sponge body. Porocytes are absent in Scypha and the ostia are simple gaps in the pinacocyte layer.

ii. Choanocytes:
(Greek: Choane, tunnel): Choanocytes (Fig. 1.25I) are also called collar cells or gastral cells, having long whip-like flagellum. Each choanocyte has a round or oval body. It possesses a single nucleus, one or more vacuoles, food vacuoles and reserve food particles.
The free end of the cell body has a comparatively longer flagellum and the base of the flagellum is surrounded by a contractile transparent collar-like outgrowth of the cytoplasm. The collars have filtering mechanisms for trapping and engulfing food particles.
Electron microscopic studies (Fig. 1.26) reveal that the collar is not a solid structure, but composed of many delicate, linear, pseudopodia-like microvilli that pass from one microvillus to the next.

The beating of the flagella pulls water into the collar. Food particles at the base of the collar are ingested as food vacuoles. The mechanism for coordinating the beat of the flagella of the choanocytes is still unknown. However, each seems to work independently. The position of choanocytes and the beating of their flagella help to direct the water towards ex-current apopyles.
iii. Mesogleal Matrix:
This gelatinous transparent matrix lies in between the two layer of cells. It forms the bulk of the body and is neither so definite nor so firm in structure. Embedded in it are spicules and numerous amoeboid cells (Fig. 1.25).
These amoebocytes carry out all the processes not accounted for pinacocytes and choanocytes. Therefore, they show greater functional plasticity. Choanocytes may sometime transform into amoebocytes.
Depending upon the size, shape of pseudopodia and kind of inclusions, amoebocytes are of the following types:
(a) Collenocytes (Fig. 1.25H):
These are amoebocytes that develop slender, branching pseudopodia, which often unite to form syncytial units.
(b) Chromocytes (Fig. 1.25E):
These amoebocytes are pigmented with lobose pseudopodia.
(c) Trophocytes:
These are nourishing amoebocytes. They receive particles of food from the choanocytes, digest and later distribute it.
(d) Thesocytes (Fig. 1.25D):
These are storage amoebocytes that have lobose pseudopodia and are filled with food reserves.
(e) Archeocytes (Fig. 1.25G):
They are undifferentiated, embryonic cells and usually form sperm or ova. They have blunt pseudopodia and large nuclei with conspicuous nucleolus.
(f) Scleroblast (Fig. 1.25F):
Some amoebocytes secrete spicules and spongin fibres of the skeleton. When they become active in skeleton formation it is called a scleroblast. They are named differently for the kind of the skeleton they form—calcoblasts form calcareous spicules, silicoblasts form siliceous spicules and spongioblasts form spongin fibres.
(g) Desmocytes:
These are elongated cells that are found in the outer cortex. It provides fibrous strength as well as helping in cortex contraction.
(vh) Myocytes (Fig. 1.25B):
These are elongated, fusiform and highly contractile cells that are similar to the smooth muscle cells. They are arranged in sphincter-like bands around the pores. They are capable of altering the diameter of the pores and regulate the amount of fluid flowing through the body.
6. Skeletal System in Scypha:
In Scypha, like other sponges soft and gelatinous body wall is supported and protected by a skeletal system. The skeletal system is formed from one or more distinctive elements—calcareous spicules, siliceous spicules and/or spongin fibres.
Calcareous spicules, characteristic of class Calcarea, are composed of some form of calcium carbonate, either aragonite or magnesium calcite. Its formation begins within the cytoplasm of a bi-nucleate calcoblast. As it grows, the calcoblast divides to form the builder or founder cell and the thickener cell (Fig. 1.27).
Each axis of a spicule begins within a separate scleroblast. In the case of a triradiate spicule, it begins within three scleroblasts and develops eventually with three founders and three thickeners (Fig. 1.27).
The founder cell moves along the surface, apparently determining the length of the ray. The thickener on the other hand, deposits additional material on the surface. After the completion of the spicule, the founder and thickener move away and merges with the mesoglea.

Siliceous spicules are characteristic of class Hexactinellida. It is composed of silicon dioxide (glass) and have an organic core in the form of an axial filament. They are formed within a single silicoblast. If they are very large then several cells may work together to form a syncytial mass within which the spicule is formed.
Spongin fibres are organic fibres made of a proteinaceous substance. Each spongioblast develops a vacuole and within it spongin materials are deposited. They live up so that the smaller fibres are united into a single long fibre (Fig. 1.27).
Sponge identification and classification depends largely upon the skeletal details. Spicules in some, make up the major skeletal elements, while in others are more or less randomly scattered in the soft parts. Spicules and spongin fibres, however, help to strengthen the poorly inter-coordinated sponge cells. Larger skeletal spicules are called megascleres and smaller flesh spicules are microscleres.
Skeleton in Scypha:
Embedded in the mesoglial matrix of Scypha are the calcareous spicules which have a definite arrangement in respect to its canal system (Fig. 1.28).

The following types of spicules are seen in Scypha:
(i) Monaxon spicules are large, straight and one-rayed. They are arranged in a circlet around the osculum forming the oscular fringe.
(ii) Oxeote spicules are monoaxon spicules that are spear-like or club-like. It projects as dense masses from dermal cortex opposite the outer ends of the flagellated canals.
(iii) Triradiate spicules are three rayed. They are present in the flagellated canals, with one ray pointed towards the closed distal ends of the canals.
(iv) Tetra-radiate spicules are four rayed and occur along with triradiate spicules in the area surrounding the spongocoel.
7. Nutrition in Scypha:
Nutrition in Scypha is holozoic and digestion is intracellular. A few amoeboid cells contain chlorophyll and are capable of carrying out autotrophic nutrition. The presence of algae and symbiotic bacteria within the sponge body, also helps in nutrition.
The food taken by Scypha is mainly protozoan. However, they do also, feed upon other microorganisms such as bacteria, diatoms and organic debris. Silica, calcium salts and other nutrients are absorbed directly from the circulating water in their canal system.
The choanocytes are the feeding organs of Scypha. The choanocytes by the activity of flagellum create water current. The food particles in the water current adhere to the outer surface of the collar of choanocytes. The food is drawn towards the collar by the undulating movement of the flagellum.
The microvilli of the collar act as a filter for trapping and engulfing food particles by pseudopodia. The effectiveness of the choanocytes in creating water current and food capture, depends upon the overall form of the organism and especially on the structure of the pore and canal system.
Calcareous sponges like Scypha have large choanocytes, where the food is digested. Digestion takes place within food vacuoles, which are acidic first and alkaline later. Various kinds of digestive enzymes are present that digest protein, starch and fat.
The digested food is passed on to trophocytes or the wandering amoebocyte. They then distribute the digested food from cell to cell like a postman distributing mail. Some digested food may be stored in thesocytes. Undigested food is egested from collar of choanocytes into the spongocoel and expelled to the exterior by the outgoing current of water.
8. Respiration in Scypha:
Special organs of respiration is lacking in Scypha. The water stream entering into the body of Scypha provides excellent conditions for respiratory exchange. Oxygen dissolved in water enters into the body by diffusion either externally through pinacocytes or internally through choanocytes.
Scypha respires aerobically and are sensitive to low oxygen demand. Some kind of oxygen debt system operates in them. When metabolism is carried out under conditions of oxygen shortage, complex organic end products are formed. These products accumulate in the body and are oxidized back when oxygen is again available. This results in a temporary rise of oxygen uptake.
9. Excretion in Scypha:
Little is known about the process of excretion. Urea, uric acid and other nitrogenous products, common in higher animals, are unknown in sponges. Excretory waste crystals, seem to accumulate in amoebocytes but have not been chemically identified. They are freed in the ex-current canals. As a general characteristic of aquatic animals, sponges too are expected to be ammonotelic in excretion.
10. Nervous System and Response Mechanism in Scypha:
Scypha like all sponges lack nerve or sensory cells. They are unable to react to a stimulus as a whole and are also unable to coordinate their actions. However, individual cells are sensitive and can react individually. Although it may seem that they are helplessly exposed to damage, they are however, left alone by fishes and other aquatic animals as they contain spicules and also as they apparently produce irritating substances.
Sponges in general do respond to certain stimuli. Myocytes, desmocytes and pinacocytes can react slowly, reducing body surface by closing ostia and oscula. When a sharp stimulus is given at a definite point it spreads slowly and evokes response in nearby cells. For example, when the osculum is pricked with a needle it results in its closure.
This kind of conduction that takes place in the absence of nervous system is termed as neuroid conduction. Power of conductivity is very low, that is reaction to light, touch and chemicals requires one to many minutes for completion.
11. Reproduction in Scypha:
Reproduction in Scypha takes place by both sexual and asexual method:
i. Asexual Reproduction:
In Scypha, asexual reproduction occurs as the result of accidental fragmentation of the body or due to exposure to unfavorable environmental conditions or it may be a part of their normal life cycle. They reproduce asexually by budding and regeneration.
Budding is the most common type of asexual reproduction. On the sides of the parent body small buds appear that grow and differentiate into young sponges. These young sponges either detach to take up independent life or remains as new members of the colony.
Scypha also has the ability to regenerate. Pieces of their body, when kept in suitable environment, produce masses of cells that develop into young sponges. They also have the ability to replace the body parts lost by injury.
ii. Sexual Reproduction:
Scypha is a monoecious (hermaphrodite) sponge. Sexual reproduction in them involves the production of gametes that generally arise from archaeocytes or choanocytes. The transformation of an archaeocyte to an ovum takes place through growth, food storage and meiosis (at the end of growth phase).
The development of sperm also takes place from archaeocyte and choanocyte. Sponges although are hermaphrodite, cross breeding is ensured by different maturation time of sperm and ova.
The sperm cells have a long tail and are able to swim freely in the water current. They find their way to the ex-current canal system and are swept out with the water current so as to eventually reach the incurrent canal system of another sponge.
The ova are amoeboid and wander through the mesoglea. The ova may grow up in size by ingesting other cells. The sperm do not directly enter the ovum, rather they take the help of the choanocyte. When the sperm cells enter the radial canal the choanocyte nearest to the egg captures and absorbs it (Fig. 1.29). This choanocyte then discards its flagellum and collar and comes closer to the egg.

This choanocyte is named as the carrier cell. The sperm loses its tail and enters the egg. The carrier cell is ultimately absorbed by the mother sponge where early development also takes place. The larva forces its way into the radial canal and finally escapes to the exterior.
Development:
The fertilized egg or zygote undergoes holoblastic cleavage and the young embryo lies embedded in the mesenchyme near the choanocytes. The first three cleavages are vertical forming 8 pyramidal blastomeres. The next cleavage is horizontal and is shifted towards the animal pole.
In this 16 cell stage, the lower tier of 8 large blastomeres are called macromeres and the upper tier of 8 small blastomeres are called micromeres (Fig. 1.30C). The macromeres lie against the parental choanocytes, while the micromeres lie in the embryo’s animal hemisphere. The macromeres stop dividing. The micromeres continue to divide and develop flagella that project into the blastocoel.
A mouth develops at the centre of the macromeres and starts ingesting parental cells. An unusual process takes place at this time, when inversion occurs and the cells pass through the mouth to turn the embryo inside out, bringing the flagella to the outer surface (Fig. 1.30F).
The parent choanocytes form a nutritive trophic membrane around the blastula. Both the macromeres and micromeres then divide to form the amphiblastula that escapes from the parent body. It then starts swimming with the flagellated micromeres in front.

At the initiation of gastrulation the amphiblastula usually attaches itself to a substratum. Gastrulation takes place by invagination or epiboly. The micromeres invaginate and move inward, while the macromeres grow over the micromeres by epiboly.
As development proceeds, the inner micromeres produce choanocytes, archeocytes, and some amoebocytes, while the outer mega meres form pinacocytes, porocytes and scleroblasts.
The larva fixes itself to a substratum at the blastopore and the central cavity forms the beginning of the spongocoel. As the spongocoel grows, an aperture called osculum appears at the free end. The embryo becomes a simple asconoid sponge, called Olynthus (Fig. 1.31).
Adult or syconoid stage is derived by the budding of radial canals. Flagellated choanocytes shift into it and the body wall further increases in thickness by the growth of middle layer with spicules and canal system. Further branching and differentiation takes place as the colony develops. Numerous pores appear on the sides to form inhalant apertures.
